技術資料
光による害虫の物理的防除方法について(その1)
技術開発室 技術研究所 環境技術グループ
キーワード
光,害虫,物理的防除,人工光源,昆虫,行動抑制
2.昆虫概論(つづき)
2.3 昆虫の視覚
2.3.1 眼の種類と構造
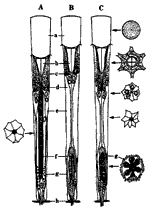
図1 コガネムシの個眼の構造(後閑,1995)
昆虫の眼は複眼と単眼に大別され,多くの昆虫は複眼を有している。図1 6)はコガネムシの複眼を構成する個眼を示し,角膜レンズ,晶子体,虹彩色素細胞,網膜色素細胞,網膜細胞,感桿(ラブドム),毛細気管,基底膜などからなりたつ。Aは昼行性の種を示し晶子体を取り巻く虹彩色素細胞や網膜色素細胞が厚くなっており,感桿は細長くなっている。Bは昼-夜行性の種の暗順応状態を示し,虹彩色素,網膜色素細胞とも薄く短く光を通し易くしている。Cは明順応状態を示し,晶子体を取り巻く2つの色素細胞は厚く長くなっている。光は網膜細胞や感桿で電気エネルギーに変換され,信号は神経細胞を経て中枢に伝えられる。また,夜間活動性の昆虫を暗所に移すと暗順応が起こり,晶子体が内方へ露出し容易に光が透入する。暗順応状態では光瞳孔が現出してくるため複眼の外観より,夜間活動性か否か判別できる。
図2 7)は単眼の構造を示し,角膜レンズ,晶子体,感桿,網膜細胞より構成されている。Aはカメムシの成虫の頭部背面にある背単眼を示し,Bはチョウの幼虫の頭部側面にある側単眼を示している。背単眼は500~1000個の視細胞からなり,レンズを1つもっている。レンズ開口が大きいため低い照度の光を受容するのに適している。側単眼はハバチの幼虫では1個,チョウの幼虫では数個頭部側方にあり,チョウではある程度のパターン認識や色の弁別も可能で,ハバチやハマキガの幼虫では偏光の検出機能があることが知られている。
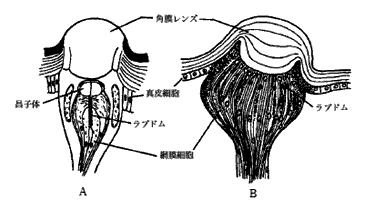
図2 単眼の構造(後閑,1995)
2.3.2 昆虫の行動
昆虫の行動様式には表5 8)に示すものがあるが,光に関連するものとしては走光性が掲げられる。走光性(Phototaxis)とは:自由運動能力を持つ生物が外部よりの光刺激に反応を起こし,この運動に方向性が認められるときに走光性と言う。走光性には積極的に光刺激源に向かうものを正の走光性,反対方向へ進むものを負の走光性と言う。さらに走光性には刺激源に進む動物の動きにより,次のようなものが知られている。
表5 昆虫の行動様式
- 無定位運動性
- 変速無定位運動性
- 変向無定位運動性
- 走性
- 刺激の種類により :
化学走性(走化性),光走性(走光性),重力走性(走地性),気流走性(走風性),水流走性(走流性),湿度走性(走湿性),温度走性(走温性),音波走性(走音性),接触走性(走触性)など - 刺激源を見分けながら進む動物の動き方により :
屈曲走性,転向走性,目標走性,保留走性(対刺激性)など
- 刺激の種類により :
- 固定的動作パターン(配偶,営巣,防御行動などの複雑な行動)
- 学習行動 : 慣れ,強化,連合(関連)学習,潜在学習
- 屈曲走光性
- 光受容器のある頭部または全身を上下左右に屈曲したり,振ったりしながら,左右から受ける光刺激の強さが等しくなるように方向を選択しながら進む。
- 転向走光性
- 光受容器が左右対称にあることを前提とし,左右の光刺激を比較し,等しい強さを受ける位置に定位して進む。光受容器を1つにすると円形運動を行いながら光刺激源に向かう。
- 保留走光性
- 網膜上に生じた像をそのままの位置に保持しようとするため,はじめに光刺激源と体の方向(体軸)との間にある角度があれば,そのままの角度を維持して進む。公園灯などの光源を目指して飛んでくる蛾は,保留走光性のため螺旋状に近づいてくる。
このような走光性が発現する最低照度はニカメイガでは 7.4×10⁻⁶~6.5×10⁻⁶ℓxと言われ,かなり低い。それ以上では,1式が成り立ち,昆虫の反応(E)は刺激光量(I)が大きければ大きいほど強くなるのが普通である。しかしながらこの反応にも限度があり,その限界を超すと走光性が逆転し,負の走光性を示すものと定位運動を停止する場合がある 9)。
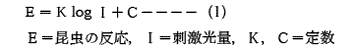
また,ニカメイガなどの走光性の波長特性として図3が良く知られているが,これは白色光及びフィルターを通した単色光(基準光源はいずれも300W 白熱電球)で両側より照射し,両者の放射照度が等しくなった点で放蛾し,両光源に向かった蛾数を数え,比刺激効果(単色光への分散蛾数/白色光への分散蛾数)としている。比刺激効果ではいずれの昆虫も紫外放射(360nm 前後)に強く誘引されることを示している 10)。
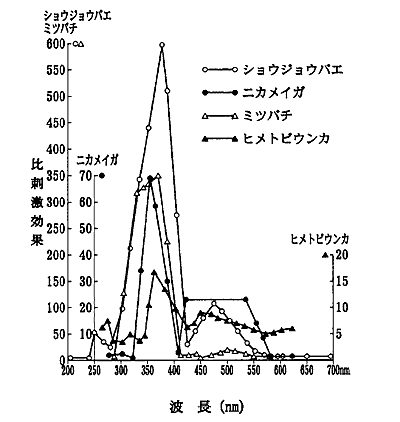
図3 ニカメイガなどの走光特性(八木,1952を改変,江村,2003)
- * ニカメイガによる加害:幼虫が稲の葉鞘や茎に,初めは集団で,後は分散して食入する。さらに出穂,登熟期の稲を食害し,鞘枯れ茎,芯枯れ茎,出すくみ穂,白穂を発生させる。
2.3.3 夜間活動性と昼間活動性
夜間活動性か昼間活動性かを,光順応の程度より評価したものを図4に示す。鱗翅目昆虫の例であるが,評点7以上は昼間活動性,7以下は夜間活動性,7前後は薄明薄暮活動性,評点下限値2以下は低照度に反応し,光源に強く誘引され,下限値3のものもかなり誘引される。下限値4~5のものは光順応能力はあるが,10ℓx以下の薄明時に活動が盛んになる。カギカッコは光順応の程度の範囲を示している 11)。また,昼間活動性の昆虫では可視放射に反応し誘引される。オンシツコナジラミは574nm 前後の冴えた黄色光に,ミナミキイロアザミウマやヒラズハナアザミウマなどは青色光に誘引される 12)。また,夜間照明において誘引される昆虫は夜間活動性のみとは限らない。昼間活動性の昆虫(トンボ,セミなど)が光による刺激を受け誘引されることは周知の事実である。
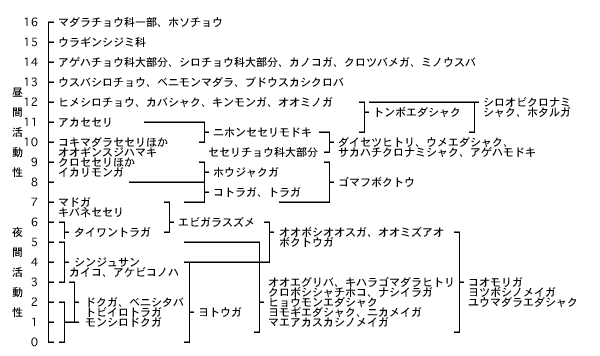
図4 複眼の構造機能による活動性の評価(河本,1992,小山,1969を改変)
2.3.4 視感度曲線
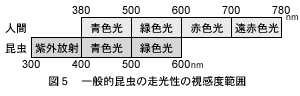
一般的昆虫の走光性の視感度は図5に示す如く,300~600nmの範囲で,人の眼の感度範囲より約100nm短波長側にシフトしている。
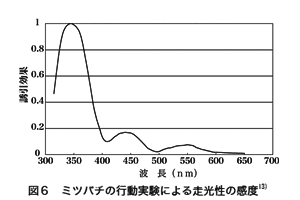
図6はどの色に良く惹かれるのかを行動学的に調べたミツバチの感度曲線である 13)。
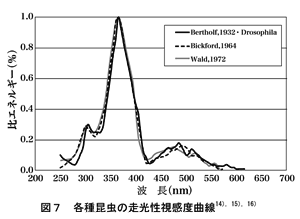
また,図7は照明関連で知られている代表的な視感度曲線を重ねたものであるが,3種の曲線は良く似ていることが判る。しかしながら,Bickford 14)の曲線とWald 15)の曲線は対象の昆虫名が不明確なことが判っている。(著者の調査による)
2.3.5 走光性の光源輝度と照度 17)
最も走光性を誘発する光源輝度は,1,000cd/m²程度とされている。光源輝度が1,500,000cd/m²程度以上になると,光源近辺に飛来後,ある一定の距離を保ち飛翔し,光源への接近飛来は行われない。走光性は数百ℓx以上の照度で抑制される。カの活動では0.1~10ℓxの照度で盛んになり,1ℓx前後が最も盛んになると言われている。
2.3.6 日週活動 17)
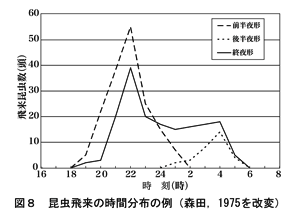
昆虫の活動(飛翔,摂食,交尾,産卵など)には周期性が認められる。昆虫は変温動物であるため,活動には一定の体温を保つ必要がある。とくに昼行性のものでは,気温,太陽放射などの影響が多く,活動の時間も規制される。図8にはガの飛来における代表的な時間分布を示す。また,カの場合は2つの活動時間帯があり,18~22時の前夜半形と朝3時~5時の早朝形となる。
参考文献
- 後閑暢夫:昆虫の生物学(第2版),玉川大出版,p.92(1995).
- 後閑暢夫:昆虫の生物学(第2版),玉川大出版,p.93(1995).
- 北野日出男:昆虫の生物学(第2版),玉川大出版,p.170(1995).
- 小山長雄:照明学会雑誌,第53巻,第3号,p.22(1969).
- 鏑木,他:照明学会雑誌,第26巻,第3号,p.84(1942).
- 河本康太郎:光バイオインダストリー 照明学会編,オーム社,p.70(1992).
- 松本義明:応用昆虫学入門,川島書店,p.100(1995).
- Helversen, O.von:Zur spektralen Unterschiedsempfindlichkeit der Honigbiene, J. comp. Physiol., Vol.80, pp.439-472(1972).
- E.D.Bickford:I.E.S Nat-conf. Paper(1964).
- Wald,G.:放射エネルギーと生命の起源-分子進化-(三浦,大井訳),東京化学同人,p.22(1972).
- Bertholf, L.M.:Z. vergleich. Physiol., Vol.18,pp.32-64(1932).
- 森田政明:照明と昆虫,松下電産技報,No.75-004,p.4(1975).
テクニカルレポートに掲載されている内容は、原稿執筆時点の情報です。ご覧の時点では内容変更や取扱い中止などが行われている可能性があるため、あらかじめご了承ください。